In this lesson, you will review how bacterial operons regulate which enzymes are expressed by genes and how this allows cells to adapt to different environments. Then, you will review how eukaryotic genes are regulated.
Escherichia coli is a bacterium that frequently lives in the human intestine and usually causes no harm. E. coli contains within its tiny structure all the genetic information required to metabolize, grow, and reproduce. It can synthesize every organic molecule it needs from nothing but glucose and some inorganic substances. Amazingly, if the organism is transplanted to a different environment, it can quickly adapt by turning off processes it no longer needs or by altering its metabolism to take advantage of new energy sources.
Here are two examples of how E. coli adapts to changing conditions:
E. coli synthesizes tryptophan from a precursor molecule in five separate steps involving five different enzymes. E. coli regulates the production of tryptophan and other amino acids through two distinct mechanisms: by direct feedback or by regulation of gene expression. The first mechanism operates all the time in an E. coli cell. When tryptophan accumulates in the cell, the activity of the enzyme in the first step of tryptophan production is repressed. The second mechanism becomes effective after a short time. If tryptophan continues to be present in the cell’s environment, the cell stops production of the enzymes in the tryptophan pathway.
The five enzymes needed by E. coli for the synthesis of tryptophan are coded by five genes that are all part of the same transcription unit. Each segment of the transcription unit is separated from the others by stop and start codons, so the single unit is able to produce five different polypeptides. A single promoter serves all five genes, resulting in a single strand of mRNA containing the codes for all five polypeptides.
The transcription unit for production of tryptophan also contains a segment of DNA called an operator. The operator is positioned within or next to the promoter. It controls the production of mRNA by allowing or blocking RNA polymerase from moving along the gene segment. The whole unit—promoter, operator, and genes—is called an operon. The operon for tryptophan production is called the trp operon from the conventional abbreviation for tryptophan, TRP.
Another gene, known as a regulatory gene, codes for a short segment of allosteric protein that acts as a repressor for this process by binding to the operator. The protein is normally the wrong shape to bind to the operator. However, when tryptophan is present, the tryptophan allosterically binds to the repressor, changing its shape and allowing it to bind to the operator. In this role, tryptophan is known as a corepressor.
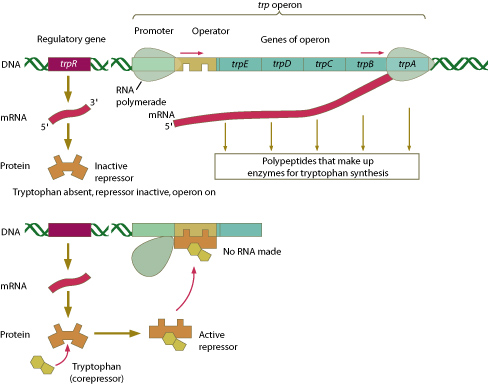
Which type of genetic sequence describes an operon?
The correct answer is A, because operons help bacterial cells adapt to changing environments. Choices B and C describe the operation of enzymes, not genes. Choice D describes the process of excretion.
The lac operon is an inducible operon. While the trp operon is repressed by environmental conditions and is called a repressible operon, other operons are stimulated by environmental conditions and are known as inducible operons.
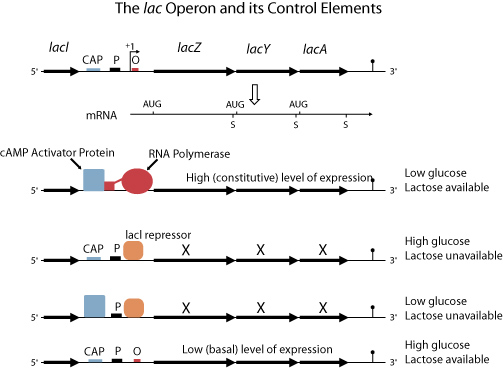
As long as glucose is present in the environment, E. coli will use glucose as its primary energy source, even when other sugars are present. However, when deprived of glucose and provided with abundant lactose, the cells of E. coli can switch over and start metabolizing lactose. There are three enzymes involved in the metabolism of lactose. The first enzyme, beta-galactosidase, separates the disaccharide lactose molecule into two simple sugars, galactose and glucose. A second enzyme, permease, codes for the membrane protein that allows lactose to pass into the cell. The role of the third enzyme, transacetylase, in lactose metabolism is unclear. It is known that transacetylase catalyzes the transfer of acetyl groups from one compound to another.
The genes for these three enzymes are contained in the lac operon, along with the promoter and operator for this gene sequence. However, there is another gene close to the operon that codes for an allosteric protein that acts as a repressor by binding to the operator in the lac operon, in effect turning it off. When lactose is present in a cell’s environment, allolactose (an isomer of lactose) allosterically binds to the repressor, changing its shape and preventing it from binding to the operator. In this way, the operon is “turned on” and starts making the mRNA needed.
Tetracycline is an antibiotic molecule that enters certain bacteria cells and allosterically attaches to a repressor protein. This prevents the repressor from binding to a gene sequence and allows a set of genes to transcribe and code for enzymes that break down the antibiotic. This set of genes is called
The correct answer is C. The gene sequence described, along with the operator and promoter, is called an operon. Choice A is the name of a gene sequence that is expressed. Choice B is the name of a gene sequence that is not expressed. Choice D is a sequence of three adjacent nucleotides that determine the insertion of a specific amino acid or signal to stop protein synthesis.

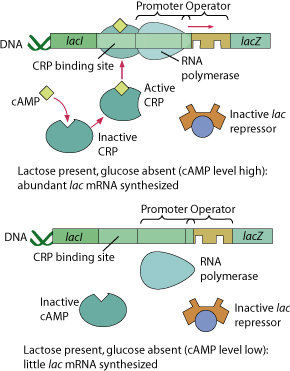
A small quantity of the three enzymes needed to metabolize lactose is always present in the E. coli cell. However, the E. coli bacterium does not use lactose as the primary energy source unless it is the only sugar present in any quantity. The E. coli cell uses another chemical mechanism to “switch on” lactose metabolism when glucose is in short supply. As in the other examples described, the mechanism depends on the interaction of an allosteric regulatory protein.
Cyclic adenosine monophosphate (also called cyclic AMP or cAMP) accumulates when glucose is absent in the environment of the cell. The allosteric regulatory protein binds to the cAMP receptors. This changes its shape, as do all allosteric proteins. The new activate shape is able to bind to another gene segment next to the lac operator and allows RNA polymerase to attach to the promoter and begin mRNA production. So there are, in effect, two “switches” that must be set properly to trigger the lac operon. One inactivates the repressor molecule and the other activates the cAMP receptor protein.
Eukaryotic cells face the same problems as prokaryotic cells. However, eukaryotic cells (except for the protists) are highly differentiated and specialized within the organism. In order to achieve this high level of specialization, different genes must be turned on or off in different cells. A highly specialized cell, such as a neuron, might express only 3–5 percent of the genes in its nucleus. A different type of cell will express a different 3–5 percent. All cells need to perform certain functions, so there are genes that are always “turned on.” However, as cells become more specialized during human development, the number of active genes in any given cell decreases.
Some genes are expressed in all eukaryotic cells at all times. These genes are responsible for the routine metabolic functions (such as respiration) common to all cells. Some genes are expressed as a cell enters a particular pathway of differentiation. Some genes are expressed all the time in only those cells that have differentiated in a particular way. For example, a plasma cell expresses continuously the genes for the antibody it synthesizes. Some genes are expressed only as conditions change around and in the cell. For example, the presence or absence of a hormone may turn on, or off, certain genes in that cell.
How many genes are estimated to be in a human cell?
C is the correct answer. Latest research shows that human cells have about 25,000 active genes. Choices A and B are too low. Choice D is too high.
There is about 1,000 times as much DNA in a human cell as in an E. coli cell, but only about 10–20 times as many genes. How can this be?
A is the correct answer. Most of the human DNA strand consists of non-coding introns. Choice B is incorrect because DNA packaging is simpler in a prokaryotic cell. Choice C is incorrect; although the genes in specialized cells may be inactivated, they are still present. Choice D is incorrect because E. coli is able to respond readily to its environment.
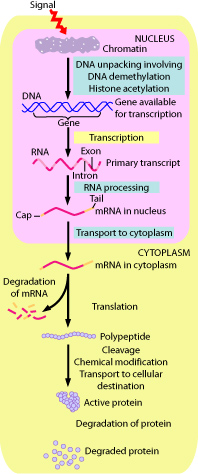
There are several methods used by eukaryotes to regulate genes. The primary method is altering the rate of transcription of the gene. Secondary methods include:
Regulation of gene transcription is the most important means by which gene expression is regulated in eukaryotes, as it was in prokaryotes. However, there are important differences between transcriptional regulation in prokaryotes and eukaryotes. Because of the complexity of eukaryotic patterns of gene expression, each eukaryotic gene is controlled by a separate promoter. Unlike prokaryotes, eukaryotic genes are not organized into operons. Prokaryotic genes are regulated primarily by repressors while eukaryotic genes are primarily regulated by transcriptional activators. These activators are called transcription factors.
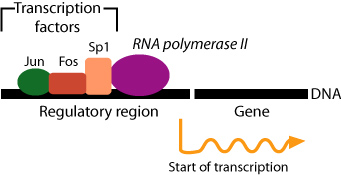
Where in the cell does gene transcription take place?
The correct answer is D, because transcription is the process of writing the code to an mRNA molecule. Choice A is incorrect; although mitochondria contain genetic information, it is separate from nuclear genetic information. Choice B is not correct because the cytoplasm just serves to transport mRNA to the ribosome. Choice C is not correct because the ribosome is the location of translation, not transcription.
Which of the following is the primary method of regulating gene expression in eukaryotes?
A is the correct answer. Regulation of gene expression is accomplished through changing the rate of transcription. Choices B, C, and D are methods of lesser importance.
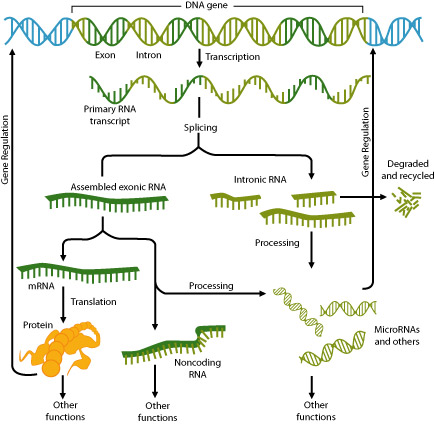
The first type of regulation can determine whether or not an mRNA gets translated. If RNA is not processed, it will not be transported out of the nucleus, and will not be translated.
The second type of regulation can affect the function of the protein produced. Some genes have exons that can be arranged in several different orders, a process called exon shuffling. When the exons are rearranged, the polypeptide produced could have a different function. For example, in mammals, the calcitonin gene produces a hormone in one cell type, and a neurotransmitter in another cell type, due to exon shuffling.
Molecules of mRNA can usually produce many copies of a polypeptide before becoming degraded. If a molecule lasts for only a short time, it will produce only a few copies of the polypeptide. A different mRNA molecule might last an hour or more, in the process producing thousands of its polypeptide molecules.
Prokaryote mRNA typically lasts for only a few minutes, which is one reason bacteria and other prokaryotes can respond so rapidly to environmental changes by varying protein synthesis. Eukaryote mRNA typically lasts much longer—hours, days, or even weeks.
The mRNAs for the hemoglobin polypeptides (alpha-globin and beta-globin) found in developing human red blood cells are unusually stable. This is the principle behind regulation of mRNA longevity. mRNAs from different genes have their approximate lifespan encoded in them. This serves to help regulate how much of each polypeptide is produced.
The information for lifespan is found in the untranslated trailer region (UTR) at the 3′ end. The sequence AUUUA, when found in the 3′ UTR, is a signal for early degradation (and therefore a short lifetime). The more times the sequence is present, the shorter the lifespan of the mRNA. Because the approximate lifetime of a strand of mRNA is encoded in the nucleotide sequence, this is a set property of each different mRNA; the longevity of an mRNA cannot be varied.
Translation of the mRNA molecule can also be regulated. Translation control mechanisms typically block the initiation stage of polypeptide synthesis. Regulatory proteins can bind to specific sequences within the leader region and the 5′ end of the mRNA strand, effectively preventing ribosomes from attaching. Global control of all or most of the translation of mRNA can be initiated by protein factors. For example, if hemes are in short supply in a developing red blood cell for some reason, a regulatory protein can inactivate a translation initiation factor by phosphorylating it. This halts all translation, but mainly affects the translation of hemoglobin mRNA since the developing red blood cell is primarily engaged in hemoglobin production.
Global control of translation is important during development of embryos. For example, many animals sequester large amounts of mRNA in their eggs, and those mRNA molecules are not translated unless the egg is fertilized. At that point, translation is triggered by the sudden activation of translation initiation factors, resulting in a burst of protein synthesis.
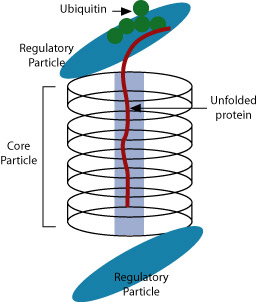
After translation, there is one more opportunity for a cell to regulate gene expression—through protein degradation. Polypeptides must be further processed to become functional proteins. For example, the protein that will become insulin must be cleaved before it is functional. Other proteins acquire sugars, and phosphate groups activate or deactivate regulatory proteins. Polypeptides frequently must be transported to other parts of the cell. All of these processes could serve as a mechanism for regulation.
Proteins marked for degradation are tagged with a small bit of protein called ubiquitin. It is not clear how proteins are chosen for destruction. Once marked, the tagged proteins are recognized by a large structure called a proteasome. The proteasome unfolds the tagged protein, sequesters it in its interior, and chops it into small peptide bits that are recycled. The ubiquitin and proteasome are also recycled.
In prokaryotes, genes that work together to produce a set of related enzymes are grouped together into an operon. Eukaryotic cells do not have operons, so they require some other mechanism for controlling several genes at the same time. This coordinated gene expression probably depends on a specific control element or set of elements that are associated with every gene in a dispersed group. A single key sets all of the genes to off or on at the same time.
In prokaryotes, an example of this sort of coordinated gene expression is the activation of a variety of genes by a steroid hormone. These steroid hormones—sex hormones are one example—have a variety of effects on the body. A steroid hormone acts like a chemical signal, entering each cell and binding to a specific receptor protein in the cell. When turned on, these receptors act like transcription activators. Every gene that will be turned on by that hormone has a control element that recognizes the transcription activator.
Genes may be turned on in one type of cell and not in another, depending on whether the transcription factor for the gene’s enhancer is active or not in a cell. For example, consider a cell in muscle tissue and a cell in skin tissue from the same person. Both have the exact same set of genes. In the muscle tissue cell, a particular set of transcription factors has become active causing the set of genes needed by muscle cells to be expressed. In the epidermal tissue cell, a different set of transcription factors is active, causing a different set of genes to be expressed.
How does the body know which transcription factors to send to which cell types and how do they get there? This is a difficult question with a very complicated answer. Many of the details have not yet been worked out by biologists. However, there are a number of mechanisms by which transcription factors can be regulated.
Steroid hormones, such as testosterone and progesterone, are derived from lipids (specifically cholesterol). Because they are derived from lipids, they can easily pass through cytoplasmic membrane into a cell, where they bind to their specific receptor. Steroid receptors are transcription factors that become activated when they bind to their ligand. Once activated, they initiate transcription of a specific set of genes.
Since peptide hormones cannot easily cross the cytoplasmic membrane, their receptors are found on the surface of the cell. When bound to its ligand, these receptors initiate a complex series of biochemical reactions inside the cell, with the ultimate result being the activation of a transcription factor often by phosphorylation, which initiates transcription of a specific set of genes.
Why are human muscle and bone cells different?
C is the correct answer. Only 2–5 percent of the genes in a specialized cell are activated. Choice A is incorrect because all of the gene sets are the same. Choice B is incorrect because eukaryotic cells are not known to contain operons. Choice D is incorrect because each specific gene produces a specific protein, unless the mRNA is damaged or reshuffled.