In this lesson, you will review evolutionary support through geology, fossils, endosymbiotic theory, the Hardy-Weinberg Equilibrium principle, and comparative anatomy of extant species.
We realize that many things change over time, such as the seasons. Yet, these do not exemplify succession through genetic inheritance as suggested by succession with modification. Central to biological evolution is the idea that all life on Earth shares a common ancestor, which gave rise to the incredible diversity evident in both the fossil record and the extant populations today.
Evolutionary change is not apparent in days, months—not even in our life time. The documentation lies within layers and layers of deposited rock accumulated over approximately 4.6 billion years. Carbon dating or radiometric dating has been used to determine the age of rocks, allowing geologists to establish geologic time divisions based changes in rock composition, as well as fossil evidence.
Geologists divide the Earth’s history into a geologic time scale that provides a framework for evolutionary theory. Based on major evolutionary events, the geologic time scale is divided by temporal boundaries marking such events. The two large divisions are called eons, which are divided further into eras.
The first eon is the Precambrian eon that spans about 4,500 million years ago (mya) to 543 mya. Precambrian is divided further into three eras. These are Hadean, Archaean, and Proterzoic.
The Hadean era spans from around 4,500–3,800 mya during which the formation of our solar system and the formation of Earth, which was uninhabitable, occurred. The Archaean era spans from about 3,850–2,500 mya and includes the formation of multiplying bacteria and photosynthesis plants. The Proterzoic era spans 2,500–543 mya and includes the stabilization of continents, build up of atmospheric oxygen, abundance of bacteria and archaeans, and presence of some eukaryotic cells.
The second eon is the Phanerozoic eon, beginning approximately 543 mya to the present. This eon is divided into three eras. The first is the Paleozoic era, spanning 543–248 mya and includes two of the most important events in Earth’s history. First, this era is marked with an explosion of multi-celled animal diversity. Second, the era ends with the largest mass extinction—90 percent of all marine species. Also, during the Paleozoic era some organisms took to a terrestrial lifestyle, while some insects took to the air.
The Mesozoic era spanned from around 65 to 248 mya. This era includes perhaps the most popular periods: Triassic, Jurassic, and Cretaceous. This was a time of dinosaurs and drastically changing fauna and flora. Amid domination of unusual plants, such as ferns, cycads, and ginkgophytes, the early gymnosperms took root, and then later the angiosperms. By the end of the Mesozoic era, non-avian dinosaurs were extinct and the diversifying angiosperms were becoming the dominant terrestrial flora.
The Cenozoic era spans from around 65 mya to present and has been typified by a diversified abundance of flowering plants, insects, fish, birds, and mammals.
Looking at the fossil record, geologists determined that there were several mass extinctions. A mass extinction is when numerous species and taxa—both marine and land—cease to exist at approximately the same time. Extinction is a continuous process; it is estimated that for every one species alive now, 2,000 have gone extinct. Background extinction is always happening at a continual low level.
It is believed that there have been about five or six mass extinctions over the history of the Earth. Based on the fossil record, there have been two major mass extinctions. The most recent was approximately 65 million years ago (mya). It marked the end of the Cretaceous period with the beginning of the end of both non-avian dinosaurs and plants. This event actually took place over millions of years—not overnight. The surviving, or extant, species then went through a period of adaptive radiation. There was a rapid increase of species diversification occurring as empty niches were gradually filled. The other major mass extinction marked the end of the Permian period (245 mya) in which it is estimated that approximately 90 percent of all marine life and a significant amount of terrestrial life went extinct.
Geologists are able to determine mass extinctions by looking at which fossils are present before and after these extinctions. For example, a large number of fossils of a certain mollusk may be found in the Permian period but not seen in any later periods. This could be of significant importance to geologists determining when the mollusk was alive and when it died out. In mass extinctions, this is seen across many species.
Extinctions are an important part of evolution. When species go extinct, their genetic complement is lost and the niches they occupied become available for other species to adapt and fill those niches.
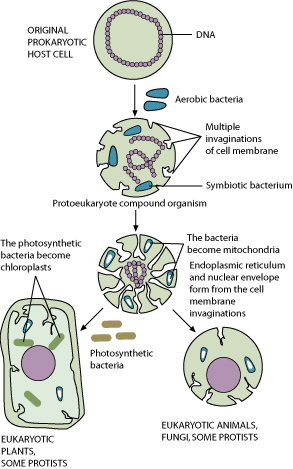
Based on the fossil record, it is believed that the mitochondrion and chloroplast organelles of eukaryotic cells evolved from free-living prokaryotic organisms. Chloroplasts were first described by Andreas Schimper in 1883, but it was not until after the discovery of mitochondrial DNA during the 1960s that the endosymbiotic theory was first proposed.
Published in 1981, Lynn Margulis’ Symbiosis in Cell Evolution included the theory that proposed that eukaryotic cells evolved from symbiotic communities of interacting elements that united together. The prokaryotes, probably chloroplasts and mitochondria, may have entered a host cell as a parasite or as ingested prey. The result may have been the evolution of a mutually beneficial interaction between prokaryote elements and the host.
The evidence for the endosymbiotic theory is remarkably solid. Mitochondria and chloroplasts:
Origin by phagocytosis is an explanation for how this actually happened. Eukaryotic cells have the ability to eat things by pulling them in and engulfing them. Once engulfed, eukaryotic cell membranes form a food vacuole around the food item and enzyme-containing lysosomes fuse with it. Typically, the item is digested. Those that do not get digested live—and may even proliferate—inside the host. Phagocytosis may also explain the double membranes of the two organelles. The original membrane could be the inner membrane, while the outer membrane originated from the food-vacuole membrane.
Although a fundamental principle in population genetics, the abstract, quantitative Hardy-Weinberg Equilibrium principle is one of the most difficult evolutionary concepts to grasp. This principle states that the genotype and gene frequencies become fixed at a particular equilibrium value after one generation of random mating. The Hardy-Weinberg principle states that the frequencies of alleles in a population do not change unless evolutionary forces act on the population. Because of this, the Hardy-Weinberg principle can be expressed as an equation that can be used to predict genotype frequencies in a population. However, this occurs only under certain conditions:
Therefore, if any of these five factors occurs, then Hardy-Weinberg Equilibrium fails. These five conditions rarely occur. If this is the case, however, then there is a formula used to determine the frequency of each allele in the population.
The basic Hardy-Weinberg formulas are:
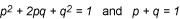
The variables are defined as:
p = frequency of the dominant allele in the population
q = frequency of the recessive allele in the population
p2 = percentage of homozygous dominant individuals
q2 = percentage of homozygous recessive individuals
2pq = percentage of heterozygous individuals
If there are only two alleles in the population, then p = 1 – q and q = 1 – p.
Alleles can be either dominant (D) or recessive (d). Genotype combinations can be DD, Dd, or dd. The Hardy-Weinberg equilibrium formula is used to determine these allele ratios in a given population and the frequencies of genotypes.
Understanding that evolution is a change in allele frequencies in the gene pool of a population, it is then the population not the individual that evolves.
There is an aspect of evolutionary biology called comparative anatomy or biology. The study compares and contrasts the structures of organisms.
Analogous structures are those with similar functions in distantly related organisms. Similar structures that are not evolutionary related are the result of convergent evolution. An example of analogous structures is the wings of insects and the wings of birds.


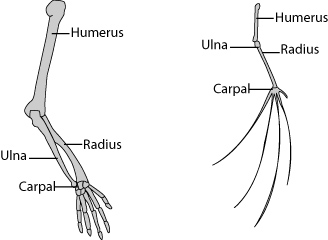
Homologous structures are those similar due to shared ancestry, yet the structures serve different functions. One form is evolutionary ancestry, in which structures evolved from a structure in a common ancestor. The arms of humans and the wings of bats is an example. Another form is developmental ancestry, in which different structures arise from the same tissue during embryonic development. The testicles of males and the ovaries of females is an example.


Although the northern gray wolf and the Mexican gray wolf are related, the Mexican gray wolf is a distinctive geographic form. The subspecies is the smallest, most genetically distinct, and southernmost occurring gray wolf on North American.
What is a species? Let us first look briefly at the scientific system of naming organisms, which revolves around the species level. For example, the scientific name, or specific epithet, of the gray wolf is Canis lupus. The scientific name is always written in italics, with the first letter uppercased for the genus name and all letters lowercased for the species name. In the case of Canis lupus baileyi, the third word is the subspecies name. A subspecies category refers to a distinctive geographic form, meaning a geographic portion of the species varies in size and/or color.
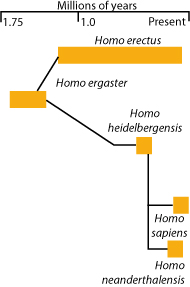
Using the fossil record, a species has been defined by its reproductive isolation from its relatives existing in a different time period. This definition has been termed chronospecies, successional species, paleospecies, or evolutionary species. An example of such a continuously changing single species would be humans: Homo erectus, H. habilis, and H. heidelbergensis.
Well before the beginning of recorded history, species have been differentiated by morphology. The term morphological species refers to a group of organisms with a distinctive form. Although this species definition is highly criticized, it is still widely used.


Another widely accepted definition for species was coined in 1942 by Ernst Mayr—a leading 20th-century evolutionary biologist. Mayr defined species as a natural population of actually or potentially interbreeding individuals that are reproductively isolated from other such groups. The definition is termed the biological species, or isolation species, concept. This proves a useful definition when working with extant examples of higher taxa that reproduce sexually, such as mammals, fish, and birds. However, it is less useful for organisms that reproduce asexually. The key to this concept is that there is no significant genetic material flow between two populations.


The phylogenetic species concept distinguishes species that share an ancestor by diagnosable geographic forms of the same basic type of organism, based on those that evolved separately. This concept works well for fossils, bacteria, and fungi. Not concerned with slightly different geographic forms interbreeding, this definition allows for different geographic groups to be separate species. This less restrictive approach would lead to many more species being designated.


No definition of species is entirely objective. Recognizing the complexity of life and the inconsistency of nature, this is no surprise—one definition rarely, if ever, can be applied across the board. Biologists may continue to debate the definition, yet most will find that it is a judgment call when selecting the most practical definition for each unique situation.