This lesson will concentrate on the theory of evolution, the foundation for all the biological sciences. We will cover the principle components and forces of evolution. As Theo Dobzhansky once said, “Nothing in biology makes sense, except in the light of evolution.”
We have reviewed classical genetics, molecular biology, and the basics of plant and animal physiology, to set the background for topics to follow.
Microevolution is a concept that refers to any evolutionary change below the level of species, the changes in gene frequency within a population or a species, and the resulting effects on the phenotype of organisms that comprise that population or species. Microevolution occurs within a short timescale, often allowing direct observation as it is happening. Processes that affect changes in a population include natural selection, mutation, genetic drift and migration.
Both Charles Darwin and Alfred Russel Wallace, independently and together, deduced the theory of evolution that has become the cornerstone of all biological sciences. Although we now have a new understanding of the genetic mechanics of evolution, the theory developed by Wallace and Darwin remains one of the most important human achievements and has undoubtedly changed the world. It is crucial to our understanding of evolution to review the basics and then come back to them in more depth.
Selection
One of the most important mechanisms of evolution is natural selection, where inheritable biological traits subject to selection are passed on to the next generation in proportion to how strongly they positively affect an organism’s survival and/or reproduction. Individuals with advantageous traits tend to be more successful reproductively than other individuals without the advantageous trait (by definition). Selection increases the frequency of these traits in a population through time, because offspring inherit traits from their parents (who produced more offspring because of the trait). Under strong selection, adaptive traits can become fixed in a population or species.
How and why selection takes place is contingent on many conditions including limited resources (e.g., food, habitat, mates), negative stressors (e.g., predators, pathogens, competition, ambient conditions), and many other factors. Natural selection is what occurs in nature, but there are other types of selection as well. Breeding of domesticated plants and animals is called artificial selection and there are ecological, directional, disruptive, sexual, and stabilizing selections that are also very important. Selection occurs when individuals in a population have at least a threshold level of variation in traits affecting survival or reproduction. Absence of variation or variation that does not impact survival or reproduction effectively precludes selection.
A considerable misconception is that selection guarantees advantageous traits will become prevalent within a population. This is not necessarily true because there are many other factors that balance evolutionary forces and effects. One of the other forces of evolution that often balances effects of selection is genetic drift. Through simple chance of genetic drift, traits that would otherwise be positively selected for can become less common or disappear entirely from a population.
Mutation
Mutations are one of the other driving forces of evolution, where deleterious mutations are usually deleted from the gene pool by natural selection, and more advantageous mutations tend to accumulate. The process of mutation involves any permanent change to DNA or RNA and is caused by many factors. Copying errors that occur during cell division, exposure to radiation, chemicals, or certain bacteria and viruses, can also occur during regular cellular processes like meiosis or mitosis. The vast majority of mutations have no real effect on the organism.
There are many types of mutations that take place and they are classified into broad groups. For example, mutations that affect one or just a few nucleotides are some of the most common. Point mutations simply exchange one nucleotide for another. Insertions augment the DNA or RNA by adding one or more extra nucleotides. Insertions often alter the proteins produced. Deletions remove one or more nucleotides and usually alter the protein that is coded by the gene.
Other mutations can affect nucleotides, chromosomal structure, and include amplifications and duplications that lead to multiple copies of chromosome regions. In contrast, deletions of large chunks of chromosomes lead to loss of multiple genes, and sometimes can affect other chromosomes as well. There are also chromosomal translocations which mix up DNA from different chromosomes, or rather chromosome inversions that switch the orientation of different parts of a chromosome and rearrange chromosomal proximity. All of these can also effectively disrupt heterozygosity with the loss of one (of a pair) allele, effectively making the new genetics homozygous. Another way to categorize mutations is by how they change the genetic functioning or the expression of the gene product.
Genetic Drift
Genetic drift is a statistical stochastic factor in biological evolution, where traits in a population (not under selection) can change over time primarily caused by randomness and chance. Basically, a random sampling of traits that do not affect reproduction or survival can become fixed or removed from a population. Because genes of every generation are not simply copied directly from the reproductively successful ancestors (they are a sample), there are statistical anomalies. Genetic drift is a cumulative effect of this sampling error on the neutral allele frequencies in the population over time. Like selection, genetic drift acts on populations, changing allele frequencies and traits. Because it is an artifact of sampling, drift is most important in small populations and can result in changes that are not adaptive.
Drift and selection
Genetic drift and natural selection are not mutually exclusive and they often occur simultaneously. Alleles are affected by drift and selection in varying ways and depending on many factors. In larger populations, for example, genetic drift would occur only slowly, and even extremely weak selection affects allele frequencies in a much stronger and faster manner. On the other hand, in smaller populations, drift can be much more pervasive. When this happens, selection may have no effect on changes in allele frequency because it is overshadowed by drift and its random sampling of alleles. At the logical extreme, in a bottleneck event where the population shrinks quickly and then regains its former numbers, genetic drift can change allele frequencies that occur independently of selection thereby eliminating many beneficial adaptations or inherent variability.
Migration
Migration (also known as gene flow) is the physical movement of genes from one population to another via individual organisms that are able to breed in a new population. Migration in or out of populations can drastically change in allele frequencies and the number of individuals with certain traits). There are a number of factors that can affect gene flow among and between populations. One of the most important is simple mobility; an individual’s mobility tends to give it greater gene flow potential.
Today we categorize four main fundamental forces that allow evolution to happen: migration, mutation, natural selection, and genetic drift. While all of these forces are important, central to this historical theory is the concept of natural selection.
Natural selection describes the process of reproductive success in organisms best adapted for their environment. Darwin and Wallace developed this concept after observing life in the tropical rain forests of the world. Darwin’s examples were brought to life dramatically by his illustrations of the finches on the Galapagos Islands. On the islands, he identified 13 species of finches. This intrigued him since he had observed only a single finch species living on mainland South America.
Darwin observed that the Galapagos finches had a wide variety of beaks and although they broadly overlapped in habitat use, they seemed to spend most of their time in completely different microhabitats. The beaks seemed adapted to different feeding habits in the diverse environmental conditions of the different microhabitats. Claw-like beaks enabled finches to eat buds and fruit. Extended beaks enabled finches to extract grubs from the ground. Heavy beaks provided the means for crushing and consuming seeds. Darwin surmised that over time, finches’ beaks evolved in order to be best suited for diverse functions, feeding on different foods. The finches with better adaptations to their environment had survived and reproduced. They had passed along their adaptations to offspring who were also more successful.
Scientists have identified many types of natural selection. Without any outside selective pressure on organisms, population traits will vary. For example, the height of adult sunflower plants varies around a mean. While most sunflower plants are around average height, others are extremely tall or extremely short. A normal bell curve reflects this trait distribution.
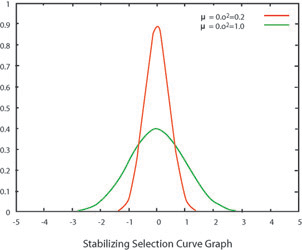
Stabilizing selection occurs when selective pressures limit variation at the extremes of the population. With the sunflower example, taller plants might not survive windy conditions, whereas shorter individuals might not survive excessive drought. In this case, the more average-height plants would become more numerous than the two extremes. As the range of traits decreases in the process of stabilizing selection, genetic diversity for that trait also decreases. Most good examples of stabilizing selection are morphological traits that seem to balance biomechanical performance and exact some sort of cost (like increased predation or caloric expenditure).
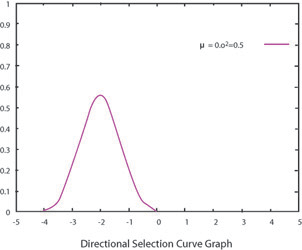
A giraffe’s long neck serves as an example of directional selection. At some time in the past, animals with short necks came under selection pressure due to competition for limited resources (leaves). After all the low leaves were eaten, animals with shorter necks were less able to reach the leafy food while animals with longer necks could reach the leaves higher up. Over time, neck length shifted in favor of individuals with longer necks because of this advantage. Therefore, one extreme of the trait distribution (shorter necks) is selected against in the process of directional selection. The trait distribution shifts to the other extreme. Just like with all forms of selection, as the range of traits decreases, genetic diversity for that trait also decreases.
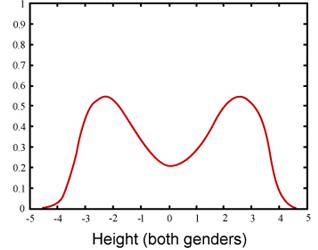
Selection pressures of disruptive selection act in the middle of the trait distribution. Let’s use an example of birds with a normal distribution of beak sizes that experiences severe declines in food availability. After a single generation, the birds in the middle of the normal curve for beak size have starved. The birds underwent extreme selection either for beaks that could fit into the tiniest nooks of a plant’s fruit to extract seeds or for beaks that were large enough to crack open nuts, for example. Those with the medium sized beaks were both unable to extract the seeds from the tiny fruits and were unable to crack open the nuts that were available to them. This is another type of selection that is theoretically possible.
Darwin had other ideas about natural selection. He thought that mate shortages produced selection pressure on species resulting in the evolution of sexually dimorphic characteristics. Previously described forms of selection have focused on the ability to survive and reproduce. Sexual selection concentrates on the mating part. Traits that improve an individual’s ability to locate, attract, mate with, or fight for access to a mate are the keys to sexual selection. Sexual dimorphism is defined as a difference in form between the sexes of a given species and is a sure sign of sexual selection.
Many species display sexual dimorphism. Male mammals are frequently larger and more colorful than females. In contrast, female insects and amphibians are often larger because they need a large body size to support egg production and carrying. Other types of sexual dimorphism include horns, antlers, eye size, coloration, and body weaponry used to acquire access to females.
The mating system of a species can affect sexual dimorphism. For primates, species that are monogamous, smaller in size, and arboreal tend to be less sexually dimorphic compared to those that are polygamous, larger in size, and terrestrial. In societal groups such as sea lions where a single male breeds with a harem of females, body and canine size are important defensive tools. On the other hand, in multi-male social groups, intelligence and communication ability are more important than sheer body size. These social groups often display cooperation to defeat dominant individuals and improve access to mating for a small group.
Sexual dimorphism, like any type of advertisement, can come with some risks. Although a brightly-colored male Northern Cardinal may attract a female quicker than dull-colored competitor, it will also have higher visibility to predators. Coloration may be a competitive advantage for mating success and a survival disadvantage at the same time. White-tailed deer provide another example. The male expends significant energy growing a rack of antlers for mating season. The male burns more calories carrying the rack throughout breeding season. Antlers provide the needed weaponry to improve access to breeding. In the bigger evolutionary picture, an individual’s reproductive success is more important than longevity. If the individual is successful in attracting, fighting for, and mating with a female, it will be passing on those adaptive characteristics to the next generation even if it only breeds for a single season.
A gene is a physical unit of heredity passed on from one generation to the next. The details of passing on genetic information happen at the allele level. An allele is one variant of a gene. Located at a specific spot on a chromosome, alleles are responsible for characteristics such as hair color, eye color, blood type, and many other inherited traits. Expressed traits represent a dominant allele while an individual may also carry an unexpressed or recessive trait.
From one generation to the next, allele diversity shifts and changes. A large, viable population of a species is able to adapt to changing conditions through genetic variability. The more alleles present, the greater the variation the building block of evolution and adaptation. Loss of genetic variation reduces a population’s sustainability. Genetic drift is the result of random sampling that is exacerbated by geographic isolation, changes in migration, and the establishment of new populations (founder effect).
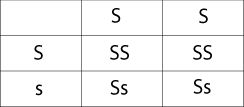
Punnett Squares can be used to graphically show how genetic variability in breeding pairs affects their offspring. The Punnett square shown below reflects breeding pairs of adults with the heterozygous gene (Ss). Offspring of these adults display genotypes (SS), (Ss) and (ss) in different proportions. The genotypes reflect all possible combinations of alleles based on the parental traits. While individual breeding pairs might not produce all of the genotypes across these percentages, a large population size could compensate for missing allele distributions.
.jpg)
What do the elephant seal and the cheetah have in common? These animals have survived the bottleneck effect. Caught in the bottleneck by hunting, habitat loss, illness, natural selection, separation through migration, and disasters such as floods, fires and earthquakes, a species may be reduced to a very small population size. The northern elephant seal and cheetah are both bottleneck effect survivors.

A small surviving population will be limited to the genetic information of its members. Genetic makeup of the survivors will be significantly less diverse than the original population. Both the northern elephant seal and cheetah reflect the DNA of genetically related individuals. While their populations have recovered from very low numbers, the limited genetic variation in current generations puts them at risk.

Wild cheetah populations currently exist in southern and eastern Africa. Genetic research indicates that cheetahs display classic characteristics of inbreeding. At one time, cheetah populations must have been reduced to very low numbers because of a genetic signature that has since been imprinted on the current populations. In order to survive, individuals bred with closely related individuals, a process called inbreeding. As animals inbreed, they pass along potentially deleterious recessive alleles that accumulate because of inbreeding. Today the worldwide population of cheetahs is nearly homogeneous genetically. The cheetah population has rebounded in size, but it is vulnerable to anything that can take advantage of this low variability, like disease, genetic defects, and sudden environmental changes.
Northern elephant seals suffer a similar plight. These animals were hunted to the brink of extinction because of their valuable reserves of blubber. The population was reduced to less than 100 individuals and these individuals became the source of genetic information for the future of the northern elephant seal. Slowly the populations began to recover and the implementation of the Marine Mammal Protection Act of 1972 enhanced the animal’s recovery potential. Today the population size may be as large as 150,000 individuals. Their limited genetic variation, however, could easily strip away all success achieved so far, and decimate the population.
As scientists observe other species that have survived the bottleneck effect they have begun to gather data about its overall impact. While species may survive and return to significantly large populations, the genetic variation remains significantly reduced. This decline in genetic variation reduces overall fitness because species have less adaptive potential. Species such as the black-footed ferret, northern elephant seal, cheetah, songbirds, whooping cranes, California condor, most whale and shark species, and many others remain at extreme genetic risk even though their numbers may be increasing.