After review of the micro-evolutionary processes, we will now turn our attention to the processes of macro-evolution, namely, speciation.
We reviewed the ideas of natural selection and the various types of selection: stabilizing, directional, and disruptive. We also covered the principles behind population bottlenecks and how they affect populations long after numerical recovery.
Macro-evolution describes the large-scale changes to an entire species. Occurring at the pace of geologic time, macro-evolutionary change may go unobserved in a human lifetime. Macro-evolution describes the loss or gain of species.
The loss or extinction of entire species has occurred throughout the course of Earth’s history. Extinction is a natural part of the ebb and flow of life. From fossil records scientists have determined that many species have lived, thrived, and ultimately become extinct. Species extinction generally occurs through a process of slow change.
Fossil records also indicate that there have been at least five periods of mass extinction. These extinctions involved the catastrophic loss of species. For example, at the end of the Cretaceous period 85% of Earth’s species disappeared. Adaptive radiation is the process of new species development following a mass extinction. Conditions created by the global change that led to the mass extinction, open the way for new ecological opportunities and creative bursts of species development.
Many factors influence the general extinction process. Natural disasters such as volcanic eruptions, earthquakes, and hurricanes can wipe out entire species. Climate change, habitat destruction, and competition between species contribute to extinction pressure. Changes in predators, parasites, and disease may also affect an entire species.
Extinction can be brought about by biological characteristics of species themselves. Species that are highly specialized, such as the koala, rely on a very specific food supply. The loss of certain eucalyptus species to deforestation has adversely affected koala populations. By comparison, coral reef species rely on precise habitat conditions. Commercial development of coastal areas has put extinction pressures on certain coral species. The corals themselves are unable to migrate or relocate to better habitat conditions. Additional risk factors include low species population numbers and slow reproductive growth capacities.
Finally, human population growth has put a significant strain on species survival. Increases in human populations have created competition over space. Construction destroys habitats needed by endangered species. At the same time it divides the space already used by some species; isolating animals and plants into small pockets making access to mates difficult. These conditions create isolation that leads to genetic drift, bottlenecks, and extinction. Humans have also begun introducing non-native species into new environments. In many cases these introduced species out-compete existing native plants and animals. Introduced species cause devastating affects on local wildlife populations. Human impact on species may be increasing extinction rates throughout the world.
Speciation is the formation of new species, an ongoing evolutionary process. New species develop out of changes in pre-existing populations. As organisms change through the forces of evolution (mutation, migration, selection, and drift), they also become unable to interbreed with their predecessors.
Speciation is one of the most important and least understood processes of biology. Although there are quite a number of definitions, the most generally used definition of species refers to groups of (actually or potentially) interbreeding natural populations that are reproductively isolated from other groups.
In scientific nomenclature, species have a unique two-part scientific name. To denote the scientific name, we either italicize or underline the genus and species names. The genus is listed first followed by the species epithet. For example, humans belong to the genus Homo, and the species sapiens, so their full scientific species name is Homo sapiens.
Speciation is the evolutionary process by which new biological species arise. There are three main types of theoretical mechanisms for the creation of new species, all based on the geographic isolation between the parent and daughter species. They range along a rough continuum from allopatric speciation (furthest) to sympatric speciation (closest), with parapatric speciation being in the middle.
Allopatric speciation happens when a population divides into two geographically isolated populations. The two different and isolated populations undergo differential divergence as they encounter and adapt to different environments. This happens because they are encountering distinct selective pressures, there is a difference in their connectivity to other populations, and/or they are independently experiencing genetic drift in different trajectories. If or when the two new species come back into contact, they have either evolved enough that they are reproductively isolated and are no longer capable of exchanging genes, or they are unable to produce viable offspring that can also interbreed.
In sympatric speciation, species diverge while remaining in close proximity and are not geographically isolated from each other. Sympatric speciation usually happens in complex biological contexts and can be found in insects that are dependent on different host plants, for example. Polyploidy is also a mechanism attributing speciation events in sympatry. Of the three different types of speciation outlined here, sympatric speciation is undoubtedly the least common and perhaps least likely to occur.
Parapatric speciation happens when the areas of two diverging populations are contiguous but do not overlap. This is only partial geographic separation, so individuals of each species may still encounter each other from time to time. This form of speciation is theoretically possible, but would take longer than pure allopatric speciation because of the potential for extended periods of interbreeding, effectively making the two populations closely related.
These types of speciation have undoubtedly taken place over the immense timescale of evolution, but it still remains a subject of debate about the relative importance of each mechanism in driving or maintaining biodiversity.
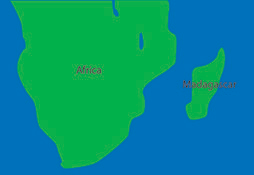
Lemurs are a primitive group of primates found on the island of Madagascar. When this island separated from mainland Africa, lemurs became isolated from other centers of primate evolution. The Madagascar lemurs now represent an endemic group of primates and they are an example of very early allopatric speciation that helped to set the stage for their subsequent adaptive radiation. Allopatric speciation occurs when individuals become geographically isolated. Over time the lemurs of Madagascar have developed adaptations making them distinctly different from the other primates in Africa. The lemurs have adapted specific traits to the diverse habitats of the island itself and have radiated to fill all potential niches of Madagascar forests.
In contrast, parapatric speciation occurs without geographic barriers for reproduction. Populations of species may share bordering territory without sharing genetic information. An example of this has been observed in an area where soil has become contaminated by heavy metals. Anathoxan odoratum is a grass species with divergent populations that have adapted by becoming tolerant to the soil’s otherwise lethal contamination. Some of the original populations of grass still grow successfully in neighboring uncontaminated soil. By growing in close range, they could fertilize one another. Through evolution, however, one population has developed an earlier flowering time than the other. Gene flow is beginning to be cut off between the tolerant and intolerant grass neighbors. This could be the start of parapatric speciation.

Evolutionary change to a species leading to the formation of another species within the same geographic area is called sympatric speciation. Sympatric speciation is a relatively rare evolutionary process in which new species develop within the range of its ancestral species. The East African Great Lakes have provided some of the few natural opportunities for researching how and why sympatric speciation occurs.
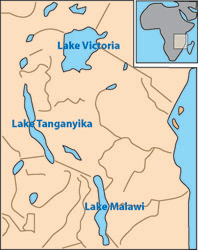
The small, deep, crater lakes in East Africa have incredibly diverse populations of cichlid fish. Sequencing the DNA of the cichlid species has revealed that many of the different species found within an individual lake are more closely related than fish from other lakes. Because the lakes are isolated from other water supplies by the natural crater rims, it is thought that cichlid species diversity developed from within. Founder individuals served as the basis for current widely diverse but genetically similar populations. The cichlids move throughout the lake encountering the diversity of microhabitats and prey items. Most species within a lake demonstrate ecological isolation in feeding styles and diet preferences ranging from open water predators to bottom detritus feeders. These ecological differences may have provided the beginning of selective mating, reproductive isolation, and finally, speciation.
Sympatric speciation is much more prevalent in plant species. Yellow bananas, cultivated wheat, rhododendron, and Appalachian spleenwort ferns are all examples of new plant species arising as a result of sympatric speciation. The acquisition of extra sets of chromosomes (polyploidy) is a major process supporting plant sympatric speciation. Sympatric speciation occurs in the range of the parent species and, as mentioned with cichlids, is a relatively rare evolutionary trajectory. Whatever the ultimate mechanism of speciation, however, there must be some type of reproductive isolation in order to maintain the separation of species’ genotypes.
In this lesson we will review the major players that helped to develop the theory of evolution, review an evolutionary timeline, and explain the gradualism and punctuated equilibrium schools of speciation. We will also review evolutionary connections determined by convergent evolution, and homologous and analogous structures.
We reviewed the main ideas behind the theory of evolution, originally proposed by Wallace and Darwin over 150 years ago. We described the four forces of evolution (mutation, migration, selection, and drift) and compared and contrasted the extremes of macro-evolution and micro-evolution. We also discussed the effects of genetic drift, population bottlenecks, and outlined two types of speciation (allopatric and sympatric).
Darwin and Wallace’s work with evolution undeniably launched a scientific revolution of research, discovery, and debate. Publishing his book, On the Origin of Species in 1859, Darwin repeatedly gave undeniably strong evidence for the concept of natural selection—first introduced in a paper he and Wallace had published nearly a year before publication of the book. Darwin and Wallace proposed the idea that those with adaptations best suited for survival and reproduction would be more successful in passing on their traits to the next generation. They knew that evolution was a gradual process in which the changes of adaptation happened slowly over many generations. According to their theory, species are always in a state of transition or continuous evolution.
At almost the same time, working with peas in a monastery garden, Gregor Mendel observed variations in plants across generations. Prior to Mendel’s work, scientists thought that a blending occurred between parents to produce the traits of the offspring. The blending passed on an “essence” to form the offspring’s new characteristics. Studying more than 28,000 plants gave Mendel the information needed to develop two important new generalizations about inheritance. Mendel’s observations led to an understanding of how inheritance worked. He discovered that discrete traits are passed along in units of inheritance. Through his research, Mendel identified the fundamental laws of inheritance and set the stage for modern genetics.
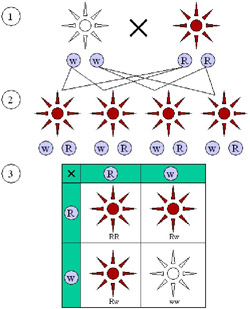
Dominant and recessive phenotypes.
(1) Parental generation. (2) F1 generation. (3) F2 generation. Dominant (red) and recessive (white) phenotypes look alike in the F1 generation and show a 3:1 ratio in the F2 generation.
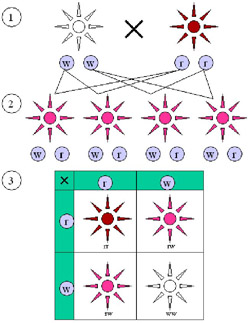
The color alleles of Mirabilis jalapa are not dominant or recessive.
(1) Parental generation. (2) F1 generation. (3) F2 generation. The “red” and “white” allele together make a “pink” phenotype, resulting in a 1:2:1 ratio of red:pink:white in the F2 generation.
Mendel’s inheritance laws changed the way scientists thought about genetic inheritance. His work was published in 1865, but it did not receive serious attention until it was re-evaluated around 1900.
Geneticist Theodosius Dobzhansky and systematist Ernst Mayr were the next key thinkers in updating ideas about evolution. In the 1930s and 1940s, they introduced a few new twists on the original theory that became known as the Modern Synthesis. Reconciling the work of both Darwin and Mendel, the Modern Synthesis shifts from a focus on individual organisms and speciation to more of an emphasis on populations, genetics, and phenotypes. The scientists argued that geographic isolation of a small sub-population would gradually lead to the evolution of new genetic traits. Through this gradualism, the changed populations become unable to interbreed with each other. Mayr clarified by saying: “Without speciation, there would be no diversification of the organic world, no adaptive radiation, and very little evolutionary progress. The species, then, is the keystone of evolution.”
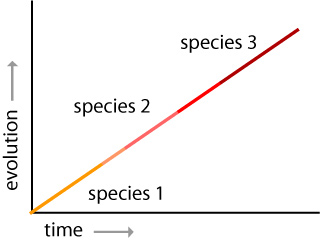
During the 1960s and 1970s, geologically trained paleontologists brought even more new research to bear on evolution. Niles Eldredge and Stephen Jay Gould reintroduced ideas about patterns of species’ stability over time. They formulated an argument around the idea of stasis where species remained in the same form with little or no change for millions of years. Eldredge and Gould thought that stasis was punctuated by bursts of rapid change when new species develop. As a result of these rapid changes, little fossil information was left behind. They called the periods of rapid change punctuated equilibrium. Eldredge and Gould demonstrated that most speciation occurred on the fringes of populations. They hypothesized that changes in species were not visible in fossil records because they occurred in very small, isolated populations. The scientists suggested that only species that were successful could leave behind enough fossil evidence to ensure a good fossil record.
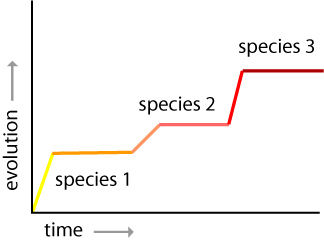
In 1985, paleontologist Elisabeth Vrba introduced the turnover pulse hypothesis. She suggested that changes in environmental conditions initiate rapid periods of extinctions and new species development. According to Vrba, external factors, such as climate changes, produce major evolutionary shifts. She studied turnover pulse events that occurred 2.5 million years ago in Eastern and Central Africa. She identified global cooling as the cause of significant African ecosystem change. Drier, cooler conditions resulted in a decline of wet woodland habitat and led to large expanses of grasslands. Species like the impala were successful in surviving the change because they were generalists. Other species were not so lucky.
Vrba also described the process of habitat tracking in which species successfully migrated into the changed habitat. She is credited with explaining that ecosystem decay and fragmentation leads to speciation and extinction.
Concurrent with all of this research has been significant breakthroughs in understanding population genetics. In 1952, Frederick Sanger uncovered the first complete protein sequence in insulin. Through his groundbreaking work, science has seen advances in DNA techniques leading to the successful conclusion of the Human Genome Project in 2001. This project set out to map the complete sequence of human chromosomes. While it seemed an impossible task, the first human DNA sequence was published in 1999 and many variations and genotypes have been added to this initial template.
Closely related to evolutionary biology and phylogeny is the study of differences and similarities between organisms, or comparative biology. Comparative biology has seen tremendous growth in the last 10 years and we now have sophisticated methods of teasing apart the evolutionary and ecological aspects that make up all life’s diversity.
Prior to Darwin’s time, Georges Cuvier was working toward significant leaps forward in our understanding of anatomy. In the late 1700s, Cuvier argued that every structure of an organism was integral to its survival. Formulating the process of comparative anatomy, Cuvier studied elephants. He compared African and Indian elephants to fossil remains of mammoths and his research led him to believe that some species that evolved between mammoths and modern elephant species became extinct. His work became the basis for modern vertebrate paleontology.
Paleontology and comparative anatomy have provided further evidence in the case for evolution. Through comparative anatomy, scientists have observed that some animals share similar structures. Homologous structures are those features shared among a set of related species. The structures are shared because they have been inherited from a common ancestor. For example, the bone structure in a human arm and a bat’s wing are homologous. While each organism has a different use for the limb (writing and flight) the general structure is similar. Scientists agree that this similarity can best be explained by common descent.
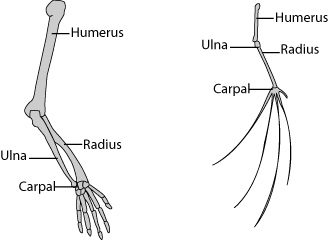
Homologous structures are the result of divergent evolution. Whales, cats, birds, bats, and humans all share this limb-structural similarity. There are many uses for the same bone structure. Locomotion strategies for swimming, flying, jumping, and writing reflect divergent evolutionary paths. Over the course of evolution then, related species become increasingly dissimilar even though they do share many traits.
Analogous structures, however, do not reflect common ancestry. Analogous structures may be superficially similar but are observed in organisms that are completely unrelated. These structures evolved in response to similar environmental factors and they evolved independently. While the structures may have similar functions, they are not the result of a common ancestor. For example, the structure of an insect wing is vastly different from a bird wing. Both will have a competitive advantage by virtue of flight. With the ability to fly an organism gains greater access to food, potential to seasonally migrate, increased access to a safe nesting space, and the ability to flee predators.
Analogous structures are an example of convergent evolution that ultimately is affected by similar selective pressures in similar ecological contexts. Unrelated species have converged upon a common feature that probably accrues some competitive advantage or an adaptation that affords some success in their environment. Dolphins and sharks are good examples, where convergent evolution has shaped their bodies in extremely similar ways. Although they are not at all closely related (one is a fish and the other a mammal), they are both strongly affected by their marine environment and the selective pressures placed on their body plans is virtually the same. Another good example is the major split within mammals — the placentals and the marsupials. Although they have lived on different continents separated for more than 55 million years, they have evolved many broad similarities. For example, the groundhog (a placental) has a strong resemblance to the wombat (a marsupial). Additionally, there are both placental and marsupial anteaters, mice, top predators, and moles. With very similar body characteristics, the main difference between these groups of mammals is how they rear their young. The marsupial’s young are born much less developed than placental young. Marsupial young develop to maturity in an external pouch. The placental mammals carry their young internally much longer. When the placental young are born, they still require significant parental care, though they are more mature than are marsupial young.